| Biodiversity at the ARC
Long-term Ecological Research Site |
Authors: Bahr, M., Bowden, B., Gough, L, Hershey, A. Hobbie, J.E, Kling, G.W., McDonald, O'Brien, W.J., Rublee, P., Shaver, G., and Walker, M.
Abstract
| Biodiversity at the ARC
Long-term Ecological Research Site |
DIVERSITY RESEARCH AT THE ARCTIC LTER SITE: A SUMMARY
Terrestrial Research on Biodiversity
The vascular flora of the Arctic is quite small in relation to the vast area that it covers, with less than 2000 species found in an area of more than 7 million square kilometers. Yet, within this small flora considerable and often striking diversity exists at the level of plant growth forms or plant functional types. Within a single watershed or toposequence, adjacent patches of vegetation may be dominated by deciduous or evergreen shrubs, by grasses or sedges, or by mosses and/or lichens. There is also considerable functional diversity within some arctic vegetation types, such as moist tussock tundra, which consists of roughly equal amounts of deciduous, evergreen, and graminoid species (Figure 1).
Functional differences among the diverse arctic plant forms have been the subject of much research over the past decades, with the result that we can now predict with a high degree of reliability the distribution of different kinds of plants within the Arctic; overall the striking functional diversity of arctic plants is tightly correlated with an equally striking microenvironmental diversity and with microenvironmental variation in general along topographic and other gradients.
What are the implications of this functional diversity at the species level for variation in biogeochemistry among arctic ecosystems? Does variation in species composition of the vegetation have any detectable effect on variation in element turnover, productivity, and organic matter accumulation in arctic landscapes, or are differences in element cycles among ecosystems controlled more by resource availability and the physical environment than by the particular species composition of a given site? These are some of the questions now being addressed in research at the Arctic LTER site (Shaver and Chapin 1991, Rastetter and Shaver 1994, Shaver 1995, Shaver et al. in press, Nadelhoffer et al. in review, Shaver et al. 1996).
One way to address these questions is by comparisons of biogeochemical characteristics among ecosystems that differ sharply in their species composition (Shaver et al.1991, Giblin et al. 1991, Shaver and Chapin 1991, Nadelhoffer et al. 1991). For example, production:biomass ratio is often used as an indication of overall element turnover in vegetation. Past research has also indicated that different plant functional types in the Arctic should differ in their production:biomass relationships. Thus, at a given level of resources, vegetations dominated by different plant functional types should differ in their production:biomass ratios and biomass turnover. However, when one actually plots production against biomass for a wide range of arctic ecosystems that differ dramatically in species composition, one finds that production:biomass relationships are nearly constant across a range in biomass of over 3 orders of magnitude (Figure 2; Shaver et al. 1996). The conclusions we draw from this comparison are as follows: first, across the full range of arctic ecosystem types, productivity and production:biomass relationships at the whole-vegetation level are controlled mainly by resource availability and environment, not by the characteristics of the individual species that make up the vegetation. Second, at any specific level of production or biomass, differences in production:biomass relationships can be detected and explained as the result of differences in turnover pattems of dominant species. These differences can be quantified as the residuals of a linear relationship between production and biomass that is determined mainly by very large differences in resource availability within the arctic landscape.
We have also used a similar analysis to detect the effects of a change in species composition on ecosystem-level C02 exchange (Figure 3; Shaver et al. in press). In this case we use data obtained from long-term (6 years) experimental manipulation of wet sedge tundra at Toolik Lake including warming, shade, and fertilizer treatments. In the treated plots, measurements of ecosystem photosynthesis were made in 1994, followed by harvesting for biomass determination, species composition, and analysis of canopy N and P content. In this experiment, ecosystem photosynthesis and canopy N mass showed a significant linear correlation despite large changes in species composition among treatments. Analysis of residuals of this relationship, however, showed that the large negative residuals for some fertilized and warmed+fertilized plots occurred in the same plots where there was a dramatic increase in abundance of Carex cordirhiza. The large negative residuals for these plots were likely due to the significantly lower photosynthetic rate of C. cordirhiza relative to the other dominant species of this site (Figure 4).
We have examined our long-term experimental manipulations of tundra to determine effects of fertilization, warming, and shading on vascular plant species richness and diversity (Figure 5, Chapin et al. 1995, Chapin and Shaver 1996, Gough and Shaver in prep.). Of the four plant communities in which we are conducting our LTER experiments, three have been harvested in recent years and were examined for both productivity and diversity responses to treatments: moist tussock tundra, dry heath tundra, and wet sedge tundra. In both the tussock and heath plots, increasing productivity by adding nutrients decreased plant species richness. One or two species increased dominance while rare species, mostly forbs, were lost. In wet sedge tundra, a four-fold increase in aboveground productivity did not affect plant species richness, although abundances of several species shifted. Warming alone did not affect species richness or productivity at the wet sedge site, but decreased species richness after 9 years of treatment at the tussock site. Shading tended to decrease both productivity and species richness.
Finally, we are attempting to better quantify patterns of vascular plant species diversity in several landscapes of different ages based on different times of glacial retreat. Within these landscapes, three communities are the subject of focus: tussock, watertrack, and snowbed. Data collection began in summer 1997 to begin to understand the effects of landscape age, soil pH, spatial heterogeneity, spatial scale, and disturbance on vascular plant species diversity. The data includes species-area relationships, dominance-diversity curves, and productivity and biomass data from a peak season harvest for both vascular and nonvascular plants. We plan to continue and expand our sampling in subsequent summers to further our understanding of controls over plant species diversity in arctic tundra.
A different approach to plant biodiversity has been taken by Walker et al. (1994) who described plant communities at two sites within the Arctic LTER research area from 154 permanent plots (each about 80 m). In addition, permanent plots of 1 m2 were established by M. Walker in 1991 at 100 m intervals in a 1 km by 1 km grid at these two sites (Toolik Lake and Imnaviat Creek) to investigate long term changes. The location of each plant in these plots has been mapped at the subcanopy level. A species list from the Toolik Lake site is given in the Arctic LTER home page. These will be re-surveyed every six years.
Aquatic Research on Biodiversity
Algae: The species of planktonic algae in Toolik Lake have been described by H. Kling (list on Arctic LTER home page). Diatom species on surfaces in the Kuparuk River have been described by Miller et al. (1991).
Bacteria: A study of planktonic bacteria has detailed the species and types in a lake (Bahr et al., 1996). Two molecular methods were used to determine the phylogenetic affinity of culturable and non-culturable bacteria from Toolik Lake, Alaska. One technique was to grow up cultures, amplify the 16S ribosomal DNA with PCR, and sequence the DNA (800-900 bases). A second technique was to concentrate bacterial cells from 40 liters of water, freeze, lyse the cells, and extract the DNA. The DNA was amplified and sequenced in the same way.
The bacteria cultured from Toolik Lake yielded seven clones that were similar to (similarity > 0.943) 16S rRNA sequences in the ribosomal RNA database. These represented a wide variety of phyla. Physiological characteristics of these cultured species include the ability to degrade macromolecules (Cytophaga aquatilus), to cause disease in plants (Burkholderia solanacaerum), to reduce nitrate (Zoogloea ramigera), to degrade low molecular weight organic compounds (Psuedomonoas mendocina), and to metabolize relatively recalcitrant aromatics (Acinetobacter calcoaceticus). Some of these species may be associated with particles.
The bacteria concentrated from lake water and analyzed by the PCR of the DNA yielded 13 clones all identified as proteobacteria. None were similar to those cultured. Six showed close affinity to Alcaligenes eutrophus, a bacteria capable of growing on organic substrates or on hydrogen plus carbon dioxide. One form, Methylophilus methylotrophus, is an aerobic methylotroph. Another, Rhodoflexus fermentatus, is a facultatively anaerobic form able to grow by photosynthesis and aerobic respiration or by anaerobic dark-fermentation. Two of the clones were related to the SAR11 cluster, a bacterium found previously only in tropical marine waters. The physiology and ecology of this form are unknown as it is known only from DNA studies. Only one form was found in both the cultures and the concentrates.
The study is one of the first of freshwaters. We hope to develop techniques so that labeled probes from these sequences can be used with the natural bacteria to determine the dominant forms throughout the year, to determine if there a succession of species throughout the year, and even to determine the chemical role of the various species.
We conclude that the culturable bacterial flora of an arctic lake contains a variety of phyla capable of metabolizing a wide range of compounds. Non-culturable forms detected only through their DNA were mostly beta-proteobacteria and included two clones related to bacteria previously found only in tropical marine waters. Molecular methods are now suitable for detailing the diversity of naturally occurring microorganisms; in the future, they should lead to information on succession, dominance, and chemical roles of the microbes.
Zooplankton: Only 12 species of pelagic crusteaceans have been recorded from the lakes and ponds of the Toolik Lake region (O'Brien et al. 1979). Surveys (at least twice) of 38 ponds and lakes (1975-1992) reveal that the number of species observed is strongly dependent upon the depth of the water body (Figure 6). This increasing species richness with increasing lake depth (which correlates with lake area) is similar to the species area curves found for many other faunas. However, the 28 extinctions and 9 invasions found do not seem to conform to the hypothesis of MacArthur and Wilson (1967) and seem to occur irrespective of lake size. Eight species are commonly collected in the ponds and lakes of the Toolik region: Daphnia middendorffiana, Daphnia pulex, Daphnia longiremis, Bosmina longirostris, Holopedium gibberum, Heterocope septentrionalis, Diaptomus pribilofensis, and Cyclops scutifer (Figure 7). Certain species do not occur in shallow ponds (Holopedium gibberum and Daphnia longiremis) while others (D. pulex) occur only in the shallowest of ponds.
The effect of predation on the zooplankton community has been described by Hobbie et al. (1995). Toolik Lake in the mid 1970's had the most diverse species assemblage with 7 of the 8 commonly found zooplankton species. This species assemblage was a combination of large-bodied and small-bodied zooplankton species thought to be made possible by reduced fish planktivory. The reduced planktivory was hypothesized to be due to large lake trout in Toolik Lake. The lake trout were thought to exclude small, planktivorous arctic grayling and lake trout from the pelagia of Toolik Lake. However, as fishing pressure removed the large lake trout in the 1970's and 1980's and planktivory by small fish increased, two of the large-bodied zooplankton ( D. middendorffiana and H. gibberum) disappeared from the lake (Figure 8).
Only females are found in the Daphnia species. Through allozyme electrophoresis and DNA sequencing (Hagen and O'Brien, unpublished), some 30 clones have been identified. For example, D. middendorffiana clones range from 7 types in one pond to 1 type in another. Studies are continuing of the response of clones to varied environments and of their genetic relationships.
Microzooplankton diversity (rotifers and protozoans) has been documented by Rublee (1992). At least twelve species of rotifers were found as well as five genera of protozoans.
Genetic variability is being investigated by Bettez and Rublee for a short-lived rotifer species, Conochilus unicornis, in several lakes near Toolik Lake. Fragments of rDNA have been amplified and cloned and are being sequenced.
Benthic Primary Producers: Bryophytes are important in both unfertilized and enriched stream habitats on the North Slope of Alaska. One common form in the Kuparuk and other similar rivers is Schistidium (Grimmia) agassazii. During the course of a 12-year long P fertilization experiment, three additional bryophyte species (Hygrohypnum alpestre, H. ochraceum, and Fontinalis neomexicana) have clearly become dominant. Thus, we have undertaken research: 1) to assess the role of bryophytes in natural, oligotrophic conditions; 2) to determine how the introduction of three new and prolific species has altered the stream ecosystem; and 3) to compare the physiological characteristics of the "stress tolerant" and the "competitive" species to determine whether they utilize different kinetic or metabolic strategies that may explain their distributions in the river. Long-term data sets and extensive literature describing the ecology of the Kuparuk River provide a unique opportunity to clearly discern the function of bryophytes in stream ecosystems.
The research we have done so far has characterized the distribution, community structure, and simple controls on production of bryophytes in the Kuparuk River (Figure 9). This research began in 1990, seven years after the P fertilization began, when we first noticed a dramatic and unexpected increase in the abundance and diversity of aquatic bryophytes in fertilized riffles in the Kuparuk River. In 1991 and 1992 the bryophyte biomass was 4-5 times greater in the fertilized reach of the Kuparuk than in the unfertilized, control reach just upstream. This was made up by three species (Hygrohypnum alpestre, H. ochraceum, and Fontinalis neomexicana) which were restricted to the high-P reach of the river. Surveys in 1993 showed that the Hygrohypnum spp. were far more abundant in riffles than in pools and that the original source of colonizing stock for these species was likely to have been nutrient-rich, headwater springs and seeps. Production by Hygrohypnum spp. and F. neomexicana in riffles was found to be strongly P-limited. Growth was most vigorous in fast flowing water and was significantly reduced in slower flowing pools. One reason for the reduction was that epiphyte and detrital abundance was 5 times greater on both artificial and real bryophyte surfaces in pools than in riffles. These bryophytes are now the dominant sink for P in the fertilized reaches and probably constitute the largest source of primary production.
Current research questions on bryophytes include the limitations on growth and physiological adaptations that give them a competitive advantage under certain environmental conditions. Research questions about ecosystem effects include the following. Does nutrient processing differ substantially in streams that have bryophytes compared to those that do not? Does the nature or the quantity of organic matter in rivers dominated by bryophytes differ from that in rivers that are not dominated by bryophytes? If so, how do these differences affect the flow of energy and matter in stream ecosystems? Are these differences predictable based on readily measurable environmental variables?
Stream Insect Diversity: Diversity of stream insects in the arctic is influenced by trophic and habitat diversity, and overwintering conditions. Overall stream habitat diversity in tundra regions is fairly low, consisting of predominantly unshaded 1st - 4th order streams flowing over boulders, cobble and peat substrates. Trophic diversity is also low, reflecting that little leafy and woody material enters streams, thereby restricting functional groups that use these resources. Because streams freeze solid, successful overwintering requires adaptations to encasement in ice during at least one developmental stage. In temperate streams mayflies (Ephemeroptera), stoneflies (Plecoptera), caddisflies (Trichoptera), and aquatic beetles (Coleoptera) are typically species rich and often account for a high proportion of secondary production. However, these important groups lose diversity with latitude and become less important trophically, while Diptera, especially Simuliidae (black flies) and Chironomidae, increase in importance. Members of these families typically have short life cycles, usually overwinter as eggs or cold-hardy larval forms, and are able to exploit the food resources and habitats available. Because most arctic streams freeze solid for many months of the year, insects with life cycles longer than one year are subjected to greater stress because of the need to overwinter more than once, possibly in more than one life stage.
Most black flies and chironomids are collectors, which use fine particulate organic matter, an abundant resource in most tundra streams. Other functional feeding groups are poorly represented. Low diversity of predators is likely due to their relatively large size and long life cycles, which makes overwintering difficult. Low diversity of shredders reflects low availability of coarse detritus, as this group is reasonably diverse in southern Alaska where coarse detritus is abundant but overwintering conditions are similarly harsh.
Chironomids are very successful in the arctic, where they are nearly as species rich as in temperate streams (Table 1). Thus, chironomid diversity does not appear to be constrained by overwintering conditions. Most arctic stream chironomids require a full year for development, although the same species may complete several generations per year in temperate regions.
Black flies are the only abundant group of filter-feeding collectors in most Alaskan arctic streams, thus occupying an important position in terms of trophic diversity. The sixteen species recorded from the Alaskan arctic (Hershey, 1995) are comparable to the same number recorded from Nebraska, where habitat diversity is also low (Table 2). In other well-studied areas with higher habitat diversity, up to 60 species have been recorded (Table 2).
These regional comparisons suggest that stream habitat heterogeneity is positively related with species richness in black flies, and overwintering conditions are of lesser importance. In arctic streams, the role of blackflies and chironomids in the food web may be especially important to ecosystem function because most other groups are poorly represented or absent.
Arctic Char Genetic Diversity: Landlocked arctic char (Salvelinus alpinus) are found in a number of lakes of glacial origin in the Toolik Lake area. Generally these lakes are small (1 - 12 hectares), deep (> 10 m), and do not contain lake trout (Salvelinus namaycush), although in at least one larger lake these species coexist. Three features make these populations particularly interesting to study. First, all arctic char in Alaska appear to be of the same lineage, based on RFLP analyses of mtDNA but show considerable diversity within the arctic lineage. Second, the glacial history of northern Alaska is unique, providing land surfaces that are of at least three different ages with respect to glacial coverage: the Itkillik glaciation ~10,000 years bp; the Sagavanirktok glaciation ~ 100,000 years bp; and the Anaktuvak glaciation >800,000 years bp (Hamilton, 1986). Third, there are currently no anadromous populations of arctic char along the northern coast of Alaska - they have been displaced by a closely related species, dolly varden char (Salvelinus malma).
Rublee and McDonald are currently studying arctic char populations from four lakes. Three of these lakes, known as the Fog Lakes, lie close together, and two are connected by intermittent streamflow. Additionally, these lakes are located on a surface that was covered, and thus the lakes formed, by the most recent glaciation approximately 10,000 years bp. The fourth lake, Oil Lake, is located 15 km west of the Fog Lakes, but lies on an older surface, last covered by glaciers approximately 100,000 year bp (Hamilton 1986, Eisner and Colinvaux 1992). We have collected 8 to 20 fish from each of these lakes, clipped the adipose fin, and extracted and purified DNA from the tissue. The purified DNA has then been subjected to a RAPD analysis using 7 different random primers. Our preliminary analysis using the Analysis of Molecular Variance Program, AMOVA, showed considerable variation within populations, no significant differences in variability among the Fog Lake populations, and a significant difference between the Oil Lake population as compared to the Fog Lake populations. Thus, the results are consistent with a long period of genetic isolation of the populations in lakes on the different glacial surfaces, despite physical proximity. We are now in the process of rerunning our RAPDs with an improved protocol to confirm these results.
We hope to extend our fish work to include two additional species, lake trout and arctic grayling, which are both more mobile than the arctic char in this environment. Results from these species should allow us to learn more about gene flow within and across watersheds for long-lived species.
Land-water interactions: the effects of species diversity
The movement of water, nutrients, and organic matter from the tundra to small streams is greatly modified by the soil and vegetation type. This effect of plant species on transferring and transforming inorganic nutrients across terrestrial landscapes has been studied at Imnavait Creek (Marion and Everett 1989) and at the Sagavanirktok River area (Giblin et al. 1991), both near Toolik Lake in northern Alaska. These studies showed that different vegetation types differ strongly in their chemical interactions with soil water and groundwater. For example (Figure 10), the amounts of N and P transported along a 1 meter wide strip were measured along a toposequence leading down to the Sagavanirktok River and traversing tussock tundra, hilltop heath, hillslope shrub/lupine, footslope Equisetum, and wet sedge tundra (Giblin et al. 1991 summarized by Hobbie et al. 1991). Some ecosystem types, like tussock tundra and dry heath, are major sources of N to soil water. Other systems, particularly those occurring under or below late thawing snowbanks, are important N sinks and P sources to soil water. Poorly drained wet sedge tundra is a P sink with a remarkably high N mineralization rate. Unfortunately, detailed studies of the flux of water and nutrients across the riparian zone have not yet been made.
Present research at the LTER site is focusing on the fluxes of nutrients and organic matter from a small watershed (8 ha). There is evidence from this small watershed (Figure 11) that the concentrations of groundwater DOC (dissolved organic carbon) are related in part to the type of overlying vegetation (Kling 1995).
Adler, P.H. and K.C. Kim. 1986. The black flies of Pennsylvania (Simuliidae, Diptera): bionomics, taxonomy and distribution. Penn State Agric. Exp. Stn. Bull. No. 856, 87 pp.
Bahr, M., J. Hobbie and M. Sogin. 1996. Microbial diversity in an arctic lake: a freshwater SAR11 cluster. Aquatic Microbial Ecology 11: 271-277.
Boerger, H.J., H.F. Clifford and R.W. Davies. 1982. Density and microdistribution of chironomid larvae in an Alberta brown-water stream. Can. J. Zool. 60: 913-920.
Bowden, W. B., J. C. Finlay and P.E. Maloney. 1994. Long-term effects of P04 fertilization on the distribution of bryophytes in an arctic stream. Freshwater Biology 32:445-454.
Bowden, W.B., B.J. Peterson, J. Finlay, and J. Tucker. 1992. Epilithic oxygen production and consumption in a fertilized arctic stream. Hydrobiologia 240:121-131.
Chapin, F. S. III, G. R. Shaver,.A E. Giblin, K J. Nadelhoffer and J. A. Laundre. 1995. Responses of arctic tundra to experimental and observed changes in climate. Ecology 76:694-711.
Chapin, F.S.III, and G.R. Shaver. 1996. Physiological and growth responses of arctic plants to a field experiment simulating climatic change. Ecology 77: 822-840.
Eisner, W.R., and P.A. Colinvaux. 1992. Late quaternary pollen records from Oil Lake and Feniak Lake, Alaska, U.S.A. Arctic and Alpine Research 24:56-63.
Finlay, J.C. and W.B. Bowden. 1994. Controls on production of bryophytes in an arctic tundra stream. Freshwater Biology 32:455-466.
Giblin, AE., K.J. Nadelhoffer, G.R. Shaver, J.A. Laundre and A.J. McKerrow. 1991. Biogeochemical diversity along a riverside toposequence in arctic Alaska. Ecological Monographs 61:415-436.
Hagen, R.H., and W.J. O'Brien. Diversity and distribution of Daphnia clones on the Alaska North Slope (Toolik Lake Area): Results from analysis of 1994 samples. Unpublished.
Hamilton, T.D. 1986. Late cenozoic glaciation of the central Brooks Range, pp. 9-49. In: T. D. Hamilton, K.M. Reed and R.M. Thorson (eds.), Glaciation in Alaska: The Geological Record. Alaska Geological Society.
Hershey, A.E., R.W. Meritt and M.C. Miller. 1995. Trophic dynamics, diversity, and life history features of black flies (Diptera: Simulidae) in arctic Alaskan streams, pp. 283-295. In: F. S Chapin and C. Kroner (eds.), Arctic and Alpine Biodiversity. Springer-Verlag, New York.
Hershey, A.E., W.B. Bowden, L.A. Deegan, J.E. Hobbie, B.J. Peterson, G.W. Kipphut, G.W. Kling, M.A. Lock, R.W. Merritt, M.C. Miller, J.R. Vestal and J.A. Schuldt. In press. The Kuparuk River: A long term study of biological and chemical processes in an arctic river. In: A. Milner and M. Oswood (eds.), Alaskan Freshwaters. Springer-Verlag, New York.
Hobbie, J.E., B.J. Peterson, G.R. Shaver and W.J. O'Brien. 1991. The Toolik Lake Project: Terrestrial and freshwater research on change in the Arctic. Proceedings of a University of Alaska Conference, "International Conference on the Role of the Polar Regions in Global Change", June 1990, Volume II: 378-383. University of Alaska, Fairbanks.
Hobbie, J.E., L.A Deegan, B.J. Peterson, E.B. Rastetter, G.R. Shaver, G.W. Kling, W.J. O'Brien, F.S. Chapin, M.C. Miller, G.W. Kipphut, W.B. Bowden, A.E. Hershey and M.E. McDonald. 1995. Long-term measurements at the Arctic LTER site, pp. 391-409. In: T.M. Powell and J.H. Steele (eds.), Ecological Time Series. Chapman and Hall, New York.
Kling, G.W. 1995. Land-water interactions: The influence of terrestrial diversity on aquatic ecosystems, pp. 297-310. In: F. Stuart Chapin III and C.Kroner (eds.), Arctic and Alpine Biodiversity. Ecological Studies, Vol. 113, Springer-Verlag.
Lindegaard-Petersen, C. 1972. The chironomid fauna in a lowland stream in Denmark compared with other European streams. Verh. Int. Verein. Limnol. 18: 726-729.
MacArthur, R.H., and H.0. Wilson. 1967. The Theory of Island Biogeography. Princeton University Press, New Jersey.
Marion, K.M., and K.R. Everett. 1989. The effect of nutrient and water additions on element mobility through small tundra watersheds. Holarctic Ecology 12:317-323.
Merritt, R.W., D.H. Ross and B.V. Peterson.1978. Larval ecology of some lower Michigan black flies (Diptera: Simuliidae) with keys to the immature stages. Great Lakes Entomol. 4: 34-39.
Miller, M.C., G.R. Hater, P. Spatt, P. Westlake and D. Yeakel. 1986. Primary production and its control in Toolik Lake, Alaska. Archiv fuer Hydrobiologie Supplementband 74:97-131.
Nadelhoffer, K J., A.E. Giblin, G.R. Shaver and J.A. Laundre. 1991. Effects of temperature and organic matter quality on C, N, and P mineralization in soils from six arctic ecosystems. Ecology 72:242-253.
Nadelhoffer, K.J., G. Shaver, B. Fry, L. Johnson, and R. McKane. In review. 15N natural abundances and N use by tundra plants. Submitted to Oecologia.
O'Brien, W. J., C. Buchanan and J.F. Haney. 1979. Arctic zooplankton community structure: exceptions to some general rules. Arctic 32:237-247.
Pruess, K.P. and B.V. Peterson. 1987. Black flies (Diptera: Simuliidae) of Nebraska: an annotated list. J. Kansas Entomol. Soc. 60: 528-534.
Rastetter, E.B., and G.R. Shaver. 1994. Functional redundancy and process aggregation: Linking ecosystems to species, pp. 215-223. In: C.G. Jones and J.H. Lawton (eds.), Linking Species and Ecosystems. Chapman and Hall, New York.
Rublee, P.A. 1992. Community structure and bottom-up regulation of heterotrophic microplankton in Arctic LTER lakes. Hydrobiologia 240:133-141.
Shaver, G.R. 1995. Plant functional diversity and resource control of primary production in Alaskan arctic tundras, pp. 199-212. In: C. Komer and F.S. Chapin, III (eds.), Arctic and Alpine Biodiversity: Patterns, Causes, and Ecosystem Consequences. Springer-Verlag Ecological Studies Series, Vol. 113. Springer-Verlag, New York. 332 pp.
Shaver, G.R., A E. Giblin, K J. Nadelhoffer and E. B. Rastetter. 1997. Plant functional types and ecosystem change in arctic tundras. In: T. Smith, H. Shugart and I. Woodward (eds.), Plant Functional Types.
Shaver, G.R., and F.S. Chapin III. 1991. Production/biomass relationships and element cycling in contrasting arctic vegetation types. Ecological Monographs 61:1-31.
Shaver, G.R., J.A Laundre, A E. Giblin, and K. J. Nadelhoffer. 1996. Changes in vegetation biomass, primary production, and species composition along a riverside toposequence in arctic Alaska. Arctic and Alpine Research 28: 363-379.
Shaver, G.R., K.J. Nadelhoffer and A.E. Giblin. 1991. Biogeochemical diversity and element transport in a heterogeneous landscape, the North Slope of Alaska, pp. 105-126. In: M. G. Turner and R.H. Gardner (eds.), Quantitative Methods in Landscape Ecology. Springer-Verlag, New York.
Shaver, G.R., L. Johnson, J.A. Laundre, D.H. Cades, A.E. Giblin, K J. Nadelhoffer and E. B. Rastetter. In press. Biomass accumulation and C02 flux in three Alaskan wet sedge tundras: Responses to nutrients, temperature, and light. Ecology.
Smock, L.A., E. Gilinsky and D.L. Stoneburner.1985. Macroinvertebrate production in a southeastern United States blackwater stream. Ecology 66: 1491-1503.
Sommerman, K.M., R.I. Sailer and C.O. Esselbaugh. 1955. Biology of Alaskan black flies (Simuliidae, Diptera). Ecol. Monogr. 25: 345-385.
Ulfstrand, S. 1982. Life cycles of benthic insects in Lapland streams (Ephemeroptera, Plecoptera, Trichoptera, Diptera, Simuliidae). Oikos 19: 167-190.
Walker, M.O., D.A. Walker and N.A. Auerbach. 1994. Plant communities of a tussock tundra landscape in the Brooks Range Foothills, Alaska. Journal of Vegetation Science 5:843-866.
Wiley, M. 1978. The biology of some Michigan trout stream chironomids (Diptera: Chironomidae). Mich.Acad. 11: 193-209.

Figure 1. Net primary production (NPP) by vascular plants in four contrasting vegetation types at Toolik Lake (including belowground stem and rhizome growth but not including root production). In (A), total NPP is indicated by the height of the bar. Within each bar, inflorescence production is indicated by the segments with vertical lines, leaf production by the unfilled segments, apical stem growth (current-year's twigs) by the dotted segments, secondary stem growth by diagonal hatching, and rhizome growth by the filled segments. In (B) through (E), the percentage of the community total NPP that is accounted for by each of the four major vascular growth forms is indicated (Shaver and Chapin 1991).

Figure 2. Total NPP versus total biomass for arctic tundra and polar desert sites dominated by a wide range of herbaceous and woody plant forms. Linear regression of these data is highly significant (P<0.01). In general, sites with negative residuals tend to be dominated by evergreen species, while sites with positive residuals tend to be dominated by graminoids, although individual sites dominated by any particular growth form can be identified along the full range of biomass and production values (Shaver et al. 1996).

Figure 3. Ecosystem photosynthesis (GEP=gross ecosystem photosynthesis) plotted versus canopy N mass in a series of experimentally treated plots in wet sedge tundra at Toolik Lake. The overall regression is significant (P<0.05), but data points with large negative residuals at high N mass are from plots dominated by Carex cordirhiza, a species with unusually low photosynthetic rates (Figure 4). (G.R. Shaver et al. unpublished)

Figure 4. Leaf photosynthesis of Carex cordirhiza and Eriophorum angustifolium, two dominant species in wet sedge tundra at Toolik Lake (Shaver et al. unpublished).

Figure 5. Vascular plant dominance-diversity curves for moist tussock tundra at Toolik Lake, sampled 3 and 9 years after initiation of experimental manipulations. Nutrient=fertilizer treatment, Temperature=greenhouse treatment, NutxTemp=fertilizer plus greenhouse treatment, and Light=shade treatment (from Chapin et al. 1995).

Figure 6. The number of species of pelagic crustacean zooplankton found in lakes and ponds near Toolik Lake, Alaska from 1975 to 1992. This data from 38 ponds and lakes are plotted against the maximum depth (meters) of each water body.

Figure 7. The percentage of ponds and lakes containing a given species of crustacean zooplankton. The ponds and lakes are divided into five categories of maximum depth. Data are from 38 lakes and ponds near Toolik Lake, Alaska, sampled from 1975 to 1992 by J. O'Brien.

Figure 8. Large-bodied zooplankton abundance in Toolik Lake, 1976-1988 (data from W. J. O'Brien).

Figure 9. Abundance (percent cover) by various bryophyte and macroalgal species in Oksrukuyik Creek in 1993. Phosphorus was added to the river at 0 km. Each bar is the mean of 5 transects at a station; the error bar is one standard deviation. This was only the third year in which Oksrukuyik Creek had been fertilized. Nevertheless, some of the same trends seen in the Kuparuk River appear to be developing in the Oksrukuyik Creek.
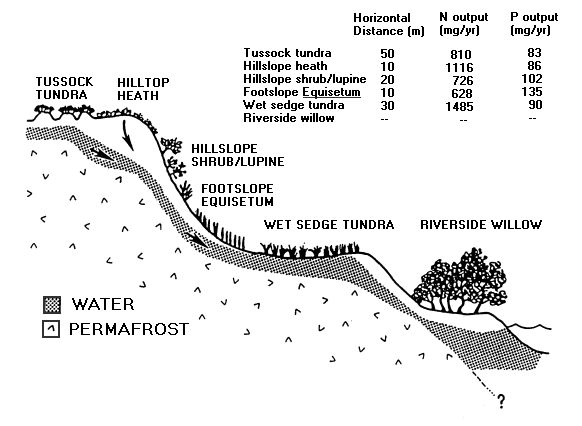
Figure 10. Ecosystem sites along a toposequence (a 1-m-wide strip) leading down to an arctic river and the amounts of N and P (mg ecosystem-1 yr-1) transported through each ecosystem annually.

Figure 11. Groundwater concentrations of DOC in a small catchment (8 ha) located near Toolik Lake, under predominantly birch, willow, or tussock vegetation.
Table 1. Comparison of species richness within the Chironomidae in selected arctic and temperate streams.
|
Stream |
Latitude |
Habitat types |
no. species |
Reference |
|
|
|
|
|
|
|
Kuparuk River, Alaska USA |
68°N |
tundra; cobble, boulder, and peat substrates |
>27 |
Hershey et al. 1995 |
|
Imnavait Creek, Alaska, |
68°N |
tundra; peat and cobble |
>40 |
Miller et al. 1986 |
|
USA |
|
substrates |
|
|
|
Caribou-Poker Creek, Alaska, USA |
65°N |
boreal forest, diverse habitats |
>35 |
Miller et al. 1986 |
|
Linding A. Western Jutland, Denmark |
56°N |
cobbles, gravel, sand, and mud substrates with dense vegetation |
64 |
Lindegaard-Petersen 1972 |
|
Hunt Creek; Michigan, USA |
45°N |
sand, clay, silt, and woody debris substrates |
>55 |
Wlley 1978 |
|
North Fork of Bigoray River, Alberta, Canada |
54°N |
diverse habitats with high macrophyte density |
32 |
Boerger et al. 1982 |
|
Cedar Creek, South Carolina, USA |
32°N |
2nd order backwater stream; snags and sand substrates |
27 |
Smock et al. 1985 |
Table 2. Comparison of black by species richness in arctic, subarctic, and temperate streams.
|
Location |
Latitude |
Habitats |
Area (km2) |
Estimated # species |
Reference |
||
|
Toolik Lake area arctic Alaska, USA |
67°N |
Tundra streams, peat and rocky substrates; trailing vegetation |
|
16 |
Hershey et al. 1995 |
||
|
Southern Alaska |
60-66°N |
Diverse mountain and boreal forest streams. |
310,800 |
38 |
Sommerman et al. l985 |
||
|
Nebraska, USA |
40-43°N |
Sand bottom channelized streams; trailing vegetation |
198,495 |
16 |
Pruess and Peterson 1987 |
||
|
Michigan, USA |
42-47°N |
Lower peninsula low gradient, deciduous forest streams on glacial moraine outwash terrain Upper peninsula boreal forest (with some northern hardwoods) streams of bedrock with low to moderate relief and including outwash/lacustrine plains. |
250,738 |
50-55 |
Merritt et al.1978; Merritt unpublished |
||
|
South Carolina, USA |
32-35°N |
Sandy, silt mud-bottomed streams at lower elevations. Swift, rocky-bottomed streams at high elevations |
78,236 |
50 |
P. Adler, pers. comm. |
||
|
Mono Co., California, USA |
38°N |
Diverse stream habitats at various altitudes with predominantly rock-bottoms. Some silt-bottom meadow streams with high concentrations of organic matter. |
8037 |
55-60 |
P. Adler, pers. comm. |
||
|
Pennsylvania, USA |
40-42°N |
Slower moving sandy-bottom streams at low elevations; swift and clear rocky-bottom streams at high elevations |
116,270 |
45 |
Adler and Kim 1986 |
||
|
Ammantls, central Swedish Lapland |
66°N |
Boreal or mixed hardwood forest streams with rocky bottoms. Birch forests at tree line |
approx. km along 3 stream reaches |
13 |
Ulfstrand 1968 |
||